一、自噬简介
1、大自噬(macroautophagy) , 也就是通常说的自噬(autophagy) , 是真核细胞蛋白降解的途径之一 。 自噬可以被描述为细胞质内的成分(细胞器、蛋白等)被双层膜的囊泡包裹 , 形成自噬体(autophagosome) , 进而传递到溶酶体进行降解的过程 。
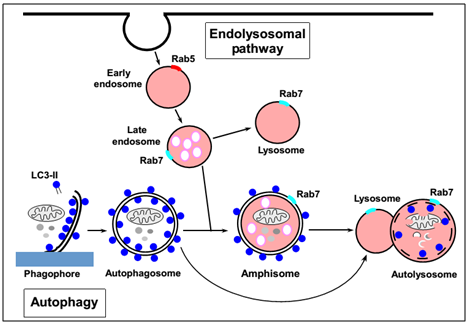
详细来说 , 自噬过程与内涵体途径(endolysosomal pathway)密不可分(见图1) 。 一方面 , 自噬体能够与晚期内体(late endosome)融合形成中间囊泡(amphisome)最终形成自噬溶酶体(autolysosome);另一方面 , 自噬体能够直接与溶酶体(lysosome)融合形成自噬溶酶体 。 无论通过哪条途径 , 自噬溶酶体最终通过酸性水解酶将细胞器、蛋白等消化分解 。
细胞本底水平的自噬发生在营养充足的条件下 , 可保护细胞免受错误折叠蛋白或受损细胞器的影响 , 从而防止某些疾病的发生(如神经退行性疾病和癌症) 。 饥饿等也可诱导自噬的发生 , 通过降解大分子物质和细胞器为细胞活动提供营养和能量 。
文章图片
图1 自噬过程及其与内涵体途径的关系
(Tom Egil Hansen and Terje Johansen , BMC Biology , 2011)
2、自噬的调控
雷帕霉素靶蛋白(mTOR)激酶在自噬反应中起着重要的调节作用 。 mTOR(Akt and MAPK signaling)在被mTOR激酶激活后 , 自噬反应被抑制;而在mTOR(AMPK and p53 signaling)未被抑制后 , 自噬反应机制被启动 。 作为酵母中Atg1的同系物 , 三个相关的丝氨酸/苏氨酸激酶 UNC-51样酶-1 , -2 , -3(ULK1, ULK2, UKL3) , 通过与mTOR复合物之间的相互作用来实现自噬反应 。 ULK1、ULK2、mAtg13和骨架蛋白FIP200(酵母Atg17的同系物)一起形成一个复杂的复合物 。 Class III PI3K复合物由hVps34, Beclin-1 (酵母 Atg6的同系物), p150 (酵母Vps15同系物), 和类Atg14蛋白 (Atg14L or Barkor) 或者紫外线放射抗性相关蛋白UVRAG (ultraviolet irradiation resistance-associated gene )蛋白组成 , 其在诱导自噬反应中起关键作用 。 Atg基因通过形成Atg12-Atg5 and LC3-II (Atg8-II)复合物来调控自噬体的形成 。 在Atg7和Atg710(分别对应泛素活化酶E1和泛素结合酶E2酶)的作用下 , Atg12和Atg5之间通过一个类泛素反应结合形成Atg12-Atg5 , Atg12-Atg5再通过非共价键与Atg16结合形成多聚体Atg12-Atg5-Atg16 , 参与自噬体的扩张 。 在Atg4、 Atg7和Atg3的作用下 , LC3转变为LC3-II-PE并紧密结合于自噬泡膜表面 , 参与前自噬体的延伸 。
自噬既能抑制也能促进细胞凋亡 , 两种反应在生物体内广泛存在 。 在营养缺乏时 , 自噬反应作为一个细胞的促活反应机制存在 , 但是过量的自噬反应也会导致细胞死亡 , 但由自噬导致的细胞死亡与细胞凋亡在形态学上有明显的不同 。 几种促凋亡信号因子 , 如TNF, TRAIL, and FADD也会诱导自噬的反应发生 。 此外 , Bcl-2也是自噬反应重要的调节因子 , 其为凋亡抑制蛋白 , 通过与Beclin-1结合形成复合物 , 来抑制由Beclin-1诱导的细胞自噬 。
线粒体自噬属于选择性自噬 , 其主要作用是降解细胞中损伤以及不需要的线粒体 。 当线粒体损伤之后 , 在正常的线粒体中持续被降解的PINK蛋白(PARL促进此反应)会处于稳定状态 , 再通过E3连接酶Parkin的作用来诱导线粒体自噬 。 Parkin会诱导线粒体膜蛋白的聚泛素化 , 由此导致与LC3结合的自噬受体蛋白的SQSTM1/p62, NBR1, and Ambra1聚集 。 此外 , 在特定的细胞类型中 , 同样存在LC3结合区域(LIR)的BNIP3 和 BNIP3L/NIX直接通过泛素依赖型反应机制也会聚集自噬反应的相关因子 , 进而促进自噬体的形成 。

文章图片
3、LC3
LC3(light chain 3)简称于MAP1LC3 (microtubule-associated proteins light chain 3) , 是目前公认的自噬标记物 。 哺乳动物中的LC3与酵母中的自噬相关蛋白Apg8/Aut7/Atg8具有同源性 。
LC3蛋白合成后在其羧基端被Atg4剪切 , 暴露甘氨酸残基 , 产生细胞浆定位的LC3-I 。 在自噬过程中 , LC3-I会被包括Atg7和Atg3在内的泛素样体系修饰和加工 , 与磷脂酰乙醇胺(PE)共价结合 , 形成LC3-II并定位于自噬体膜上(见图3) 。 哺乳动物中的LC3可分为三种: LC3A、LC3B和LC3C 。其中 , LC3B作为LC3的一种 , 同样可以用作自噬的分子标志 。
【细胞质|知识分享:自噬研究】
文章图片
图3 LC3在自噬体形成中的作用
(Varuna C. Banduseela et al , Phyciological Genomics , 2012)
4、自噬流检测方法
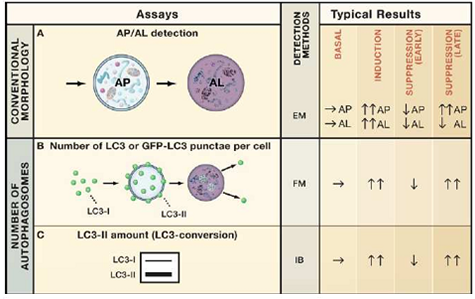
细胞经自噬诱导或抑制后 , 需对自噬流的水平进行观察和检测 , 常用的技术手段见表1:
文章图片
表1 自噬体计数及自噬流检测方法汇总
(Mizushima et al, Cell, 2010)
其中 , 电镜观察法由于受到实验条件和设备的限制应用范围有限;而由于自噬过程发生快速 , 通过IB/WB检测LC3-II水平和GFP-LC3的方法易受到实验进行时长的影响 。
想要准确检测自噬流的变化 , 需要多种技术手段共同使用 , 才能达到理想的实验效果 , 产生具有说服性的实验结果 。
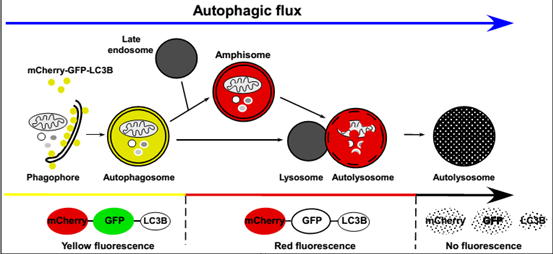
自噬双标产品 :mCherry-GFP-LC3B腺病毒(Adenovirus)、慢病毒(Lentivirus)和腺相关病毒(AAV)
mCherry-GFP-LC3B自噬双标系统的工作原理为: 未发生自噬的细胞及含有自噬体的细胞中 , 由于mCherry与GFP共同表达 , 细胞呈现黄色荧光 。 当自噬体与溶酶体融合形成自噬溶酶体后 , 酸性的溶酶体环境使酸敏感的GFP荧光淬灭 , 而mCherry不受影响 , 进而使自噬溶酶体呈现红色荧光 。 因此 , 红色荧光可指示自噬溶酶体形成的顺利程度 。 红色荧光越多 , 绿色荧光越少 , 则从自噬体到自噬溶酶体阶段流通得越顺畅 。 反之 , 自噬体和溶酶体融合被抑制 , 自噬溶酶体进程受阻(见图4和附图) 。
维真现推出mCherry-GFP-LC3B腺病毒、慢病毒和腺相关病毒 , 满足您实验的不同需要 。
·简便直观 , 实现自噬可视化;
·准确指示 , 实现自噬流监测 。
文章图片
推荐阅读







- 新机|快准稳,捕捉每个高光瞬间! 刘衡分享EOS R5拍摄体验
- 问答|紧追B站加码知识类内容,抖音上线“学习频道”
- 显示器|华光昱能知识--DP2.0跟DP1.4有那些不一样?
- 审判|直接服务“三城一区”主平台,怀柔科学城知识产权巡回审判庭成立
- 词条|百度百科上线2500万词条,超750万用户参与共创科普知识内容
- 风险|筛选风险词、用实体类型推断限售商品,阿里安全夺冠知识图谱大赛
- 字节跳动|抖音上线学习频道,为知识内容增加一级入口
- 知识科普|直管内径、凸缘管内径密封快速接头 管内壁粗糙的管口格雷希尔GripSeal连接器
- 训练|华为运动健康 Beta 新版本测试:新增健身课程分享、血压挑战计划
- 声音|魏晨分享视频感谢粉丝,手机录视频无修音唱歌,瘦到面部凹陷